| N-T.ru / Раритетные издания / Пётр Маковецкий |
Задача 108-4. Спортлото и жизнь на других планетахПётр Маковецкий. Смотри в корень! Сборник любопытных задач и вопросов А.Читая научную литературу, иногда встречаешь утверждения, вызывающие острое желание испытать их на прочность. Это можно сделать многими путями. Один из них – проверка предпосылок, следствием которых является утверждение. Другой – извлечение следствий из самого утверждения и их анализ на правдоподобие. Если они оказываются правдоподобными, то это увеличивает шансы на правильность утверждения, в противном случае – ставит его под сомнение. Вот одно из утверждений, достаточно далеко идущих и поэтому интригующих. Н. Рашевский вычислил общее количество различных биологических видов Ν, существование которых, по его мнению, принципиально возможно*. По этим расчетам, N = 108. Это число первым путем проверить трудно: мы недостаточно компетентны в биологических предпосылках и математических методах, которыми оно получено. Пойдем вторым путем: будем извлекать следствия, считая N = 108 правильным, и посмотрим, что из этого получится. * N. Rashevsky «Topology and Life», «Bulletin of Mathematical Biophysics», 1954, v. 16, p. 317; краткое изложение – в книге И.А. Акчурина «Единство естественнонаучного знания», «Наука», 1974, стр. 108. На Земле в данное время имеется около 2 млн видов* (1,5 млн видов животных и 0,5 млн – растений), т.е. в 50 раз меньше, чем их может быть вообще. Если же учесть, что многие виды существовали раньше, но вымерли, и на всю длительность эволюции накинуть еще 2 млн видов, то всего на Земле уже реализовано n = 4·106 из N = 108 возможных. Н. Рашевский выдвигает смелую гипотезу, что биологическая система (в дальнейшем, для краткости, – биология) другой планеты может состоять из других видов, входящих в названное число N. Мы принимаем и эту мысль, потому что она правдоподобна и, кроме того, что уж совсем ненаучно, она нам нравится. * «Общая биология», учебник для 9...10 классов, под ред. Ю.И. Полянского, «Просвещение», 1973, стр. 10. Давайте, однако, решим такую задачу. Поскольку, по-видимому, жизнь на планетах у разных звезд возникала независимо, то, скорее всего, многие виды у обеих биологий неодинаковы, но есть шанс, что некоторые из видов совпадут. Будем считать, что обе биологии обладают одинаковой мощностью n = 4·106, и найдем вероятность того, что вторая биология перекрывается с земной хотя бы в одном из видов. Сведения по теории вероятностей, которые нам понадобятся, настолько элементарны, что они найдутся практически у каждого из вас. Б.Событие A: «Две биологии перекрываются хотя бы в одном из видов» – очень сложное, оно распадается на множество более простых: Таблица 6
и т.д. вплоть до полного совпадения по всем видам. Подсчитать W(A) – вероятность события A – значит подсчитать ее для всех взаимоисключающих вариантов Ai и затем сложить. Это громоздко. Проще вычислить вероятность противоположного события A и потом, воспользовавшись тем, что прямое и противоположное события составляют полную группу событий A и A, для которой W(A) + W(A) = 1, найти W(A) из формулы
Итак, W(A) есть вероятность того, что ни один из видов биологии-2 не совпадает с соответствующим видом биологии-1 (земной). Задача несколько прояснится, если мы предварительно решим похожую, другую, более житейскую и с меньшим числом возможных исходов. Как известно, в карточке спортлото всего N = 49 номеров, подчеркнуть нужно n = 6 номеров (чем больше угадаешь, тем больше выигрыш), В данном случае W(A) есть вероятность того, что вы угадаете хотя бы один номер (т.е. или один, или два, ..., или шесть). Здесь противоположное событие W(A) есть вероятность того, что вы не угадаете ни одного номера. Аналогия будет понятнее, если мы будем считать, что тираж уже состоялся, 6 некоторых номеров уже выиграли, отмечены в тиражной таблице, но нам результаты еще не известны (угадывать до тиража принято только во избежание злоупотреблений). Земная биология есть таблица на 108 номеров, по которой «тираж» уже состоялся: 4·106 номеров (видов) уже выиграли, остальные – нет. Биология-2 «подчеркивает» свои номера, ничего не зная о том, какие номера выиграли у нас. В.Итак, в таблице спортлото подчеркнуты (или будут подчеркнуты) неизвестные нам 6 номеров из 49. Подчеркивая в своем билете наугад одну из цифр, мы имеем 6 шансов из 49 угадать, а остальные 49 – 6 = 43 – не угадать. Вероятность не угадать при первой попытке W1(A) = 43/49. Вероятность не угадать ни одного номера в двух попытках
где W2(A/1) – вероятность не угадать во второй попытке при условии, что мы не угадали в первой. Легко видеть, что W2(A/1) = (48 – 6)/48 = 42/48. В самом деле, теперь мы подчеркиваем один из 48 номеров (а не 49): не будем же мы еще раз подчеркивать уже подчеркнутый номер; в числителе вычитается по-прежнему 6, потому что мы исходим из условия, что не угадали в первый раз (условие 1, см. выше): W2(A) = (43/49)·(42/48). Аналогично W3(A) = (43/49)·(42/48)·(41/47). И, наконец.
Итак в спортлото шансы не угадать вообще ни одного из шести номеров довольно велики: 44 из 100. Очевидно, вероятность угадать хотя бы один номер W(A) = 1 – W(A) = 1 – 0,44 = 0,56. Для упорядочения записи домножим числитель и знаменатель формулы (3) на два факториала: W6(A) = (43·42·41·40·39·38)/(49·48·47·46·45·44)·(43!/43!)·(37!/37!). После очевидных упрощений W6(A) = (43!43!)/(49!37!) = [(N – n)!(N – n)!]/N!(N – 2n)!. Теперь перейдем к биологии. Здесь вместо шести сомножителей-дробей в формуле (3) их будет четыре миллиона: W(A) = (108 – 4·106)/108·(108 – 4·106 – 1)/ или в записи через факториалы:
Вычислить непосредственно это число вручную – непосильный труд. Для обхода этой трудности существует приближенная формула Стирлинга:
Относительная ошибка здесь меньше, чем второе слагаемое в скобке, которое при m = 108; 96·106; 92·106 составляет около одной миллиардной от всего результата. В конечном итоге ошибка будет еще меньше, так как после подстановки (5) в (4) скобки числителя и знаменателя почти точно сокращаются. Поэтому второе слагаемое скобки мы можем отбросить. Кроме того, сокращаются (точно) e и √[2π] и остается W(A) = {[(96·106)^[96·106]·(96·106)^[96·106]]/ Разделив числитель и знаменатель на (106)^[192·106], получим W(A) = (96^[96·106]·96^[96·106])/(100^[100·106]·92^[92·106])·(96/[10√[92]]) = Теперь окончательный результат нетрудно получить логарифмированием: lgW(A) = 92·106(2lg96 – 2 – lg92) + 8·106(lg96 – 2) + lg96 – 1 – (1/2)lg92 ≈ Таким образом: W(A) ≈ 10–68000. Остается преодолеть одну вычислительную трудность: поскольку мы пользовались четырехзначной таблицей логарифмов и в скобке вычитались друг из друга близкие числа, то относительная ошибка разности логарифмов существенно превосходит относительную ошибку самих логарифмов. В частности, мы не можем в числе 0,0008 поручиться даже за один-единственный знак, который там есть. Следовало бы взять, например, семизначную таблицу логарифмов, которой, однако, у автора под рукой не оказалось*. Но в этой задаче есть возможность поступить иначе. Можно обойтись оценкой числа W(A) сверху, т.е. таким числом, которое не слишком отличается от W(A) и в то же время достоверно больше его. Вероятность W(A) будет наверняка больше истинной, если вместо 0,0008 мы возьмем 0,001 (наверняка большее истинного) и вместо 0,0177 возьмем 0,016 (наверняка меньшее). Тогда * Не надо считать автора лентяем: найти семизначную таблицу было бы значительно проще, чем суметь обойтись без нее не в ущерб задаче. Оценка сверху – тоже один из приемов при вычислениях. Его тоже полезно принять на вооружение. lg W(A) < 92·106·0,001 – 8 – 106·0,016 = – 36 000. Теперь мы можем утверждать, что W(A) < 10–36000. Обратите внимание на то, что мы отважно идем на преувеличение W(A) в 10–36000/10–68000 = 1032000 раз и тем не менее получаем результат, выводы из которого останутся* теми же. * Кстати сказать, более точный результат (с помощью цифровой вычислительной машины, заменившей десятичную таблицу логарифмов) еще более впечатляющ: W(A) < 10–72404. Что это за выводы? Главный: вероятность полного несовпадения двух биологий невообразимо мала. Слово «невообразимо» здесь не для красного словца: число 10–36000 действительно невообразимо. Это число с полным спокойствием и чистой совестью перед нашей конкретной задачей можно считать нулем. И, следовательно, согласно формуле (1) W(A) = 1 , т.е. факт перекрытия двух независимых биологий хотя бы в одном из видов – достоверен*. (Должно быть не менее 1036000 биологий, чтобы хотя бы один случай полного несовпадения стал реальным. Но ведь во всей видимой Вселенной звезд всего лишь 1021. Значит, для одного случая полного несовпадения нужно иметь 1035979 Вселенных (!!), причем имеющих биологию у каждой звезды.) * Для любителей математических тонкостей достоверности рекомендую книгу Э. Бореля «Вероятность и достоверность», Физматгиз, 1961. Из достоверности этого факта следует парадоксальный вывод: все биологии похожи друг на друга! Ведь совпадение в одном из видов (неважно в каком, пусть даже в вымершем!) делает биологии практически родственными, так как каждый вид родствен всем (или большинству) видам своей биологии, а его двойник в другой биологии – всем видам своей (мы исходим из гипотезы о единстве происхождения всех или хотя бы большинства видов внутри данной биологии*)). Это, в частности, сильно увеличивает шансы на наличие других цивилизаций и на возможность контакта с ними. * На Земле бактерии и сине-зеленые водоросли считаются неродственными остальной биологии (животным + растениям). См. стр. 71 упомянутого учебника биологии. Интересно подсчитать, сколько в среднем видов в двух биологиях оказываются совпадающими. Пусть наша биология является мишенью (по форме напоминающей таблицу спортлото), содержащей N = 108 квадратиков, из которых n = 4·106 зачернены. Биология-2 делает n = 4·106 «выстрелов» по этой мишени. Стреляет куда попало, но не мимо таблицы (это означало бы, что в биологии-2 существуют виды, не входящие в число 108). Сколько в среднем будет попаданий в черные квадратики? Поскольку черные квадратики составляют m = n/Ν = 1/25 часть всей мишени и стрельба ведется беспорядочно, то 1/25 часть всех выстрелов придется на черные квадратики, т.е. число совпадающих видов M составляет 1/25 от всех существующих в данной биологии: M = nm = n2/N = (4·106)2/108 = 160 000. Это число может вызвать некоторые сомнения. Для уточнения следовало бы учесть, что при беспорядочной стрельбе возможны и двойные (и тройные) попадания в один квадратик, что в стрельбе естественно, а в биологии не имеет смысла: это означало бы, что «стреляющая» биология уже внутри самой себя содержит два (или три) совпадающих вида. Но совпадающие виды – это один вид, т.е. совпадающие результаты надо считать за один выстрел. Поэтому более подходящей моделью является такая мишень, из которой после каждого выстрела изымается пораженный квадратик (черный или белый), а следующий выстрел обязательно попадает в один из оставшихся квадратиков. Однако эта уточненная модель даст то же самое. Ведь из-за двойных и тройных попаданий в отдельные квадратики некоторые другие квадратики не получат причитающейся им пули. Поэтому после окончания стрельбы (4·106 выстрелов) M’ будет меньше вычисленного M. Но если подсчитать «излишки» пуль, полученных некоторыми квадратиками, и произвести соответствующее дополнительное число выстрелов (повторяя, если надо, эту процедуру несколько раз), то в пределе мы как раз и получим M = 160 000. Впрочем, M’ ненамного меньше М: вероятность двойного попадания в один квадратик равна m2 = 1/625, тройного – m3 и т.д. Число совпадающих видов M = 160 000 является наиболее вероятным, отклонения от него возможны в обе стороны (теоретически вплоть до M = 0 и до M = n), однако с помощью теории вероятностей можно показать, что отклонение от M на 10√[M] практически невозможно: вероятность такого события порядка 10–50. Итак, практически достоверно, что M = 160 000 ± 4000. 160 000 совпадающих видов! 160 000 прямых «родственных связей» между двумя биологиями! А еще надо учесть, что остальные, несовпадающие виды косвенно родственны совпадающим! Можно ли в такой ситуации ожидать, что вторая биология существенно отличается от первой, земной? Можно сделать и еще один, самый сенсационный, пожалуй, вывод. Поскольку средняя плотность видов равна n/Ν, то такова же должна быть в среднем и вероятность совпадения для каждого конкретного вида, в том числе и человека! Пусть α – событие «совпадение человек – человек (а остальные – как угодно)». Тогда W(α) = n/N = 1/25. Это очень большая вероятность. А вероятность возникновения цивилизации кажется еще большей: ведь для этого не обязательно совпадение «человек – человек»; возможно, соседний человеку вид (в нашей биологии отсутствующий) тоже может создать цивилизацию*. Но это число не должно нас обольщать: оно основано на средней плотности видов и для одного конкретного вида крайне ненадежно. Вокруг некоторых из видов пустых мест может оказаться гораздо больше: вполне возможно, что в некоторой биологии будет провал не только в окрестностях вида «человек», но даже отряда «приматы», класса «млекопитающие», а возможно, провал охватывает даже весь тип позвоночных. Возможно даже, что позвоночные – аномалия (на языке теории вероятностей – грубый промах) биологии, т.е. в других биологиях они встречаются крайне редко. Наконец, возможно, что биология – вообще аномалия природы. С другой стороны, цивилизацию, возможно, могли бы построить потомки осьминогов. * Вероятность встречи двух цивилизаций может оказаться намного меньше вероятности возникновения каждой из них: по некоторым пессимистическим оценкам (которых автор не разделяет, но обойти молчанием не может) срок существования цивилизаций ограничен, цивилизации смертны. Это значит, что две цивилизации могут не встретиться из-за несовпадения во времени существования (аналогия – задача о встрече X и Y, см. ниже). Вероятность W(α) = 1/25 может оказаться существенно ошибочной для одной пары биологий, но если взять большое число биологий, то для них в среднем эта W(α) будет верной. Таким образом, если даже всего биологий в Галактике порядка тысячи, то вероятность того, что еще какая-нибудь из них, кроме нашей, содержит человека, становится практически равной единице. Фантасты приучили нас к тому, что на других планетах мы встретим нечто совершенно непохожее на земную биологию. Эта привычка очень сильна в нас и заставляет усомниться в правильности наших выводов. Нет ли в них ошибки? А если есть, то где она? Она может быть либо у Н. Рашевского (возможно, число потенциально допустимых видов значительно больше 108), либо в наших расчетах, либо в нашей попытке перенести модель спортлото на биологию. Научная этика требует: прежде, чем утверждать, что ошибся предшественник, искать ошибку у себя. Свои расчеты автор проверил многократно, ошибки не нашел (может быть, найдет читатель?). То, что мы предполагали одинаковую мощность множеств обеих биологий (4·106), не влияет на вывод о достоверности. Вывод будет тем же, если мощности не равны, а только сравнимы по порядку величины. Рассмотрим внимательнее модель спортлото. Соответствует ли она биологии? Каждый номер в спортлото выпадает почти независимо от других пяти номеров. Так ли в биологии? Нет! Каждый вид – следствие эволюции некоторого предка и следствие борьбы за существование с современниками. Шесть выигрышных номеров спортлото могут оказаться рассеянными в случайном порядке по всему множеству, состоящему из 49 видов спорта; 4·106 видов биологии более или менее связаны происхождением и вряд ли будут разбросаны по всему множеству возможных 108 видов.
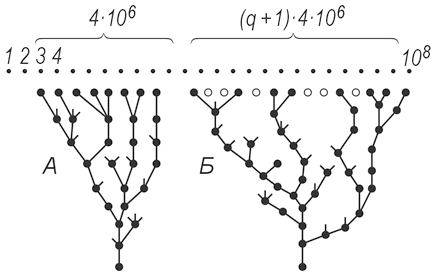
Рис. 156. Модель биологии мощностью 4·106 видов Допустим, что все 4·106 видов данной биологии представляют собой тесную группу (A на рис. 156, сравните с рис. 11 в упомянутом учебнике биологии). Тогда задача о частичном совпадении двух биологий напоминает не задачу о спортлото, а известную по многим книгам задачу о встрече. Два лица X и Y условились встретиться в определенном месте между 12 и 13 часами. Пришедший первым ждет второго 10 минут, после чего уходит. Какова вероятность встречи, если X и Y приходят независимо в случайный (в пределах указанного часа) момент?
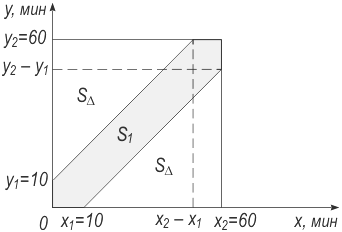
Рис. 157. Задача о встрече Обозначим момент прихода лица X через x, лица Y – через y. Встреча состоится, если |x –y| ≤ 10 мин. В декартовой системе координат область выполнения неравенства (заштрихована на рис. 157) имеет площадь S1 = Sквадр – 2SΔ = 602 – 2·(502/2) = 1100. Искомая вероятность W(A) = S1/Sквадр = {y2x2 – [(y2 – y1)(x2 – x1)/2]}/y2x2 = [N2 – (N – n)2]/N2 = 1100/602 ≈ 0,306. Применив новую модель к нашей задаче (y2 = x2 = N; y1 = x1 = n), имеем
Эта величина кажется более естественной: вероятность, что две биологии не перекрываются, равна W(A) = 1 – W(A) ≈ 0,9216. Следовательно, возможны непохожие на нас биологии. С другой стороны, если биологий во Вселенной не две, а сотни, то и вероятность перекрытия многих из них между собой довольно велика. Результат успокаивает и фантастов, и реалистов. Само собой разумеется, что если n ≥ Ν/2, то даже компактные биологии перекрываются достоверно. Они просто не могут не перекрыться: им двоим в пределах N не хватает места! В самом деле, если N = 108 и, например, n = 60·106, то
Это что еще за новость? Вероятность неперекрытия равна 1–0,84 = 0,16 ≠ 0! Как могут две биологии поместиться на отрезке N без перекрытия, если каждая из них занимает более половины этого отрезка? Что-то неладно. Хорошо, что нам пришло в голову проверить то, что «само собой разумеется». Хуже было бы, если бы ошибку обнаружили читатели. В чем же ошибка? Опять не та модель? Вернемся к задаче о встрече. Внимательный анализ ее показывает, что она действительно имеет не замеченное ранее отличие от нашей задачи. По условиям задачи лицо X (или Y) может прийти на встречу в любое мгновение между 12 и 13 часами, в том числе и в 12 ч 59 мин, и прождать условленные 10 минут. При этом 9 минут ожидания из 10 приходятся на 14-й час и не имеют смысла для увеличения вероятности встречи, но не противоречат условиям задачи. В нашей же задаче по условиям этого делать нельзя: обе биологии должны целиком принадлежать множеству 108. Для наших целей задачу о встрече нужно модифицировать. Все условия остаются теми же, кроме одного: самый поздний момент, когда партнеры обязаны явиться, – это 12 ч 50 мин, чтобы все 10 минут ожидания находились внутри 13-го часа. Модификация показана пунктиром на рис. 157. Теперь при n < Ν/2
а при n ≥ Ν/2 W(A) = 1. Итак, задача решена. Смущает одно: в нашей модели компактной биологии все 4·106 видов поставлены вплотную друг к другу, занимают смежные точки. Но если это так, го как пойдет дальнейшая эволюция? Существующим видам некуда развиваться. Правда, могут появляться новые виды на краях «генеалогического дерева» (рис. 156, A), но крайними являются лишь десятки или сотни видов (дерево нужно представлять не плоским, как на рис. 156, A, а многомерным). Поэтому основная масса видов лишена возможностей развиваться: все потенциально возможные в этой области виды уже существуют. Эволюция закончилась, прогресса больше не предвидится! В это плохо верится. Видимо, гипотеза о компактной биологии тоже неверна. По существу, два рассмотренных варианта являются двумя крайностями, истина где-то посредине. Биология более или менее компактна, если она объединена родством происхождения; в то же время между существующими видами должны быть вакантные места, чтобы развитие не остановилось. Крона «дерева» должна быть не такой густой, но зато более раскидистой (рис. 156, Б). Этого требует дивергенция (см. учебник биологии). Если вокруг каждого существующего вида имеется в среднем q вакантных мест, то вся биология (крона «дерева») раскинута в пределах (q + 1)·4·106. Какое число q близко к истине? Автор не берется ответить на этот вопрос. Допустим, что q = 9, т.е. биология мощностью в 4·106 видов раскинула крону своего «дерева» на «площади» (9 + 1)·4·106 = 40·106 из всей возможной площади в 108, Тогда, согласно (8), вероятность перекрытия биологий (не отдельных видов, а крон «деревьев»)
вероятность неперекрытия W(Б) = 1/9. Нетрудно видеть, что третья биология с такими же n и q уже никак не может быть втиснута в N без перекрытия. И, скорее всего, она будет перекрываться как с первой, так и со второй. Если же биологий во Вселенной много, то подавляющее большинство из них перекрывается со многими другими. Нужно, однако, сделать одну оговорку. Мы еще не нашли вероятность пересечения видов. Вероятность пересечения видов W(A) в данном случае есть вероятность перекрытия биологий W(Б), умноженная на вероятность пересечения видов W(A/Б) при условии, что биологии перекрываются, т.е.
(сравните с формулой (2)).
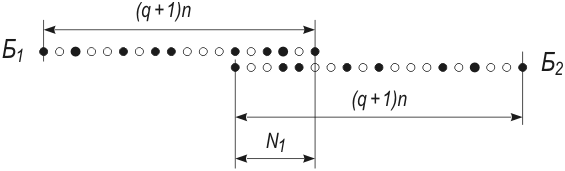
Рис. 158. Вариант перекрытия двух биологий Когда мы говорили о перекрытии двух биологий, содержащих по q вакансий на каждый вид, то не упоминали о возможности того, что одна биология может перекрываться с другой так, что существующие виды одной будут попадать на вакансии другой. Правда, поскольку вакансии данной биологии находятся в тесной близости к ее существующим видам, окружая почти каждый из них, то даже такое «перс-плетение крон деревьев» без совпадения уже должно говорить о похожести биологий. Но мы этим не удовлетворимся. Мы вычислим второй сомножитель W(A/Б). Если две биологии перекрываются хотя бы на отрезке в N1 – 5000 точек (большинство из которых – вакансии, см. рис. 158), то область перекрытия можно рассматривать как таблицу спортлото (все-таки она пригодилась!), для которой N1 = 5000 и n1 = N1/(q + 1) – число номеров, которые нужно подчеркнуть. При q = 9n1 = 500. Тогда W(A/Б) = (N1 – n1)!(N1 – n1)!/N1!(N1 – 2n1)! = Обратите внимание: Ν1 = 5000 составляет лишь одну восьмитысячную часть от каждой из биологий, т.е. в рассмотренном примере относительная ширина перекрытия ничтожно мала. И тем не менее вероятность совпадения видов уже равна W(A/Б) = 1 – (A/Б) > 1 – 10–20, т.е. совпадение видов становится таким же практически достоверным, как и при перекрытии компактных биологий (различия между вероятностями 10–20 и 10–∞ не имеют практического значения). Итак, второй сомножитель в формуле (10) равен единице, W(A) = W(Б), и поэтому формула (9) дает правильный результат, упомянутую оговорку можно не учитывать. Если q + 1 > 12,5, т.е. вакансий более 92%, то перекрытие двух любых биологий и совпадение некоторых видов обеих биологий достоверно. Поскольку, однако, эти результаты кажутся невероятными, то, видимо, следует пересмотреть в сторону увеличения число N = 108, полученное Н. Рашевским*. И тем не менее очень хочется, чтобы Н. Рашевский оказался прав: тогда биологии разных планет будут обязательно похожими. Это пригодилось бы нам для четырех следующих задач. * Недоверие к N = 108 только усилится, если учесть, что не все виды, существующие и существовавшие на Земле, уже открыты и, возможно, n >> 4·106. Более того, при конечном N конечна и эволюция, что вызывает философские возражения. * * * Данная задача может произвести на читателя странное впечатление. Автор выдвигает одну модель, рассчитывает ее, потом спохватывается, бракует модель, просчитывает новую, опять ошибается, вводит в модель изменения, опять бракует ее и, наконец, рассматривает модель, почти не отличимую от первой. При этом он то делает определенные выводы, то опровергает их и т.д. Не удивляйтесь! Все так и задумано. Изложение этой задачи – попытка показать кухню научной работы, процесс созревания идеи, муки творчества, т.е. то, что при публикации результатов всегда отсутствует. Должен сознаться, мук, зигзагов и ошибок было еще больше. Однако оставленные за бортом зигзаги и ошибки были уж настолько нелепыми, что публичная их демонстрация ничего, кроме конфуза для автора, не дала бы. Эта же задача, если бы автор решился опубликовать ее в научном журнале, выглядела бы так. К вопросу о вероятности пересечения земной и внеземной биологииН. Рашевским* на основе предложенного им принципа биотопологического эпиморфизма построен топологический комплекс-граф наиболее существенных для всякого живого существа отображений. На основании асимптотических оценок числа графов данного структурного типа он получил общее число различных биологических видов, которые принципиально могут существовать: N = 108. * N. Raschevsky «Topology and Life», «Bulletin of Mathematical Biophysics», 1954, v. 16, p. 317. Единственная известная нам биология содержит n = 4·106 видов (включая 2·106 вымерших). Полагаем вместе с Н. Рашевским, что биологии других планет могут содержать в себе и другие виды из числа N. Рассмотрим вероятность пересечения двух независимых абстрактных биологий равной мощности n, все виды которых принадлежат множеству N. Мы анализируем три модели биологий. 1. Рассредоточенная биология: все виды двух биологий рассредоточены случайным образом по множеству N (в силу дивергенции). Тогда вероятность того, что ни один из видов биологии-2 не совпадает с соответствующим видом биологии-1 (земной), равна W(A) = [(N – n)!]2/N!(N – 2n)! < 10–36000. Ничтожность этого числа означает, что противоположное событие W(A): «две биологии пересекаются (совпадают) хотя бы в одном из видов» – достоверно. Математическое ожидание числа совпадающих видов M = 160 000 со среднеквадратичным отклонением σ = 400. Но если это так и если внутри каждой из биологий все n видов (или хотя бы сравнимая с n часть) объединены родством происхождения, то между собой биологии оказываются квазиродственными через совпадающие виды, т.е. между двумя биологиями нет различий, которые можно было бы назвать существенными. В частности, это говорит в пользу того, что вероятность существования внеземных цивилизаций не является малой. 2. Компактная биология: все n видов расположены на множестве N тесной группой (вследствие единства происхождения). Тогда вероятность пересечения двух биологий W(Б) = W(A) = 1, n > N/2. Для n = 4·106 W(Б) ≈ 0,082, т.е. умеренна. Однако вероятность пересечения быстро возрастает с ростом числа биологий и при пБ > 25 становится достоверностью. 3. Промежуточная модель: биология близка к компактности (в силу единства происхождения) и в то же время имеет вакантные места (условие продолжения эволюции). Среднее число вакансий на один реализованный вид равно q. Тогда W(A) = W(Б)W(A/Б) = где W(A/Б) – вероятность совпадения хотя бы одного из видов при условии перекрытия биологий на подмножестве Ν1. Даже при очень малых Ν1 ≈ √[n] и q = 9 второе слагаемое в фигурной скобке имеет величину менее 10–20. В результате W(A) = W(Б) = 8/9. При q > 11,5 W(A) = 1. Поскольку q = 11,5 нельзя считать слишком большой величиной, то третья, наиболее близкая к реальности, модель биологии дает практически достоверное пересечение для любых двух биологий. Итак, две биологии разных планет не могут быть не похожими друг на друга. Этот вывод следует из полученного Н. Рашевским N = 108 и может оказаться неверным только при N >> 108. П.В. Маковецкий Как видите, научная статья пишется кратко (потому что она пишется для специалистов и, кроме того, необходимо экономить бумагу). При этом уже не видны творческие муки, статья кажется чудом озарения свыше, так как исчезли промежуточные выкладки и рассуждения, т.е. убрана та лестница, с помощью которой только и можно было добраться до результата. Часто это затрудняет чтение статьи даже специалисту. Правда, при этом экономится бумага, но упускается из виду, что экономить следовало бы не бумагу, а мозги читателей. Цель нашей книги несколько иная: привить читателям вкус к всестороннему рассмотрению проблемы, показать не только результат, но и путь к нему, равно как и заблуждения, подстерегающие нас в пути.
• Задача 108. Свидание под часами |
|
Дата публикации: 3 марта 2007 года |
|